Hc2
Hoorcollege
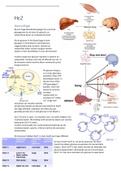
Bij een hoge bloedsuikerspiegel zijn vooral de
glycogenese (in de lever & spieren), en
vetsynthese (lever en vetweefsel) actief.
Als de glucose in het bloed laag is komt
glucagon in het bloed en wordt glucose
uitgescheiden door de lever. Spieren en
vetweefsel zullen minder op gaan nemen,
zodat er meer beschikbaar is voor de hersenen.
Insuline zorgt voor glucose-opname in spieren en
vetweefsel. Insuline remt ook de afbraak van vet. In
de hersenen werkt insuline alleen remmend op het
hongercentrum.
Als glucose hoog is,
is er meer glycolyse,
waardoor meer ATP
beschikbaar komt. In
de pancreas zitten
ATP-gevoelige K+-
kanalen. Door het
kanaal te remmen
ontstaat
repolarisatie en
ontstaat Ca2+-influx.
Ca2+ zorgt voor
exocytose van insuline-vesicles.
Het glucose-kanaal van de pancreas (GLUT-2) heeft
een lage affiniteit, waardoor de cellen dus pas
gevoelig worden als er veel glucose in het bloed is.
GLUT-4 komt in spier- en vetcellen voor. De cellen hebben een
insulinereceptor. Bij binding vindt exocytose plaats van vesicles
waarop de GLUT-4 zitten.
Insuline veroorzaakt een conformatieverandering van de
insulinereceptor, waarna interne reacties de exocytose
veroorzaken.
De hersenen hebben GLUT-3, deze heeft een hoge affiniteit.
De lever heeft GLUT-2, net als de pancreas. De lever
neemt dus alleen glucose op wanneer de concentratie
hoog is. Door GLUT-2 kan zowel secretie als absorptie van
glucose plaatsvinden, afhankelijk van de concentraties.
GLUT-2 is hier dus essentieel om glucose te exporteren.
,Insuline zorgt voor glycogeensynthese in de lever en spieren.
Glycogeensynthase en glycogeenfosforylase worden
hoofdzakelijk gereguleerd. Glycogeensynthase maakt de
lange 𝛼1,4-ketens. Een ander enzym zorgt voor de 𝛼1,6-vertakkingen. De vertakking is efficiënter.
Voor afbraak is naast fosforylase ook een debranching enzyme nodig voor de 𝛼1,6-bindingen.
Er is nooit 0 insuline of 0 glucagon, er is altijd een
bepaalde verhouding.
Om eiwitten van conformatie te laten veranderen
kun je deze fosforyleren (door kinase) of door te
binden op een allostere site.
Glucagon bindt een 7TM-receptor waardoor cAMP
in de cel vrijkomt → cAMP → fosforylatie
fosforylase kinase → fosforylatie
glycogeenfosforylase. glycogeenfosforylase kan
glycogeen afbreken.
Tegelijkertijd wordt de glycogeensynthese geremd. Hetzelfde PK-A fosforyleert glycogeensynthase.
Hier is echter juist de niet-gefosforyleerde vorm actief.
Insuline werkt tegengesteld aan glucagon. Het werkt door fosfatasen te activeren, die de
fosforyleringen ongedaan maken. Als er geen insuline is, is er geen cAMP, en overheerst de fosfatase.
De irreversibele glycolysestappen worden geregeld. Het
belangrijkste regelpunt zit bij PFK.
Fructose-2,6-BP remt fructose-1,6-bisfosfatase, en stimuleert
fosfofructokinase (PFK), hiermee stimuleert het de glycolyse.
Insuline zorgt dat fructose-2,6-BP in hoge concentratie aanwezig
is. Glucagon zorgt juist dat F-2,6-BP afgebroken wordt.
PFK wordt verder gestimuleerd door AMP & Pi, en geremd door
ATP, H+, en citraat.
Hexokinase wordt geremd door negatieve feedback van G-6-P.
Pyruvaatkinase wordt gestimuleerd door feed-forward van F-
1,6-BP, en geremd door ATP en alanine.
Na lang vasten wordt via transcriptieregulering de expressie van G6Pase, F-1,6-BPase, PEPCK, en
pyruvaatcarboxylase verhoogd, om zo de gluconeogenese te bewerkstelligen.
Cortisol stimuleert middels genexpressie de
gluconeogenese.
,Hc3
Hoorcollege
Belangrijkste lipiden: TAG, vrije vetzuren (FFAs, NEFAs (non-esterified)), fosfolipiden, en cholesterol.
Endogene lipiden zijn vrije vetzuren en TAG. De voornaamste lipogenetische cellen zijn adipocyten en
hepatocyten. Ook melkklieren kunnen vet maken. Vet wordt altijd vanuit acetyl-CoA gevormd.
Lever: TAG → VLDL in bloed, de lever slaat dus geen vet op maar transporteert het naar het bloed,
waarna het door adipocyten kan worden opgenomen. Vet kan ook TAG maken, maar slaat het op.
De basis voor vet, acetyl-CoA, komt meestal uit glucose
(en soms uit aminozuren). Acetyl-CoA uit pyruvaat wordt
in het mitochondrion gemaakt. De vetzuursynthese vindt
echter in het cytosol plaats. Via de citraatshuttle wordt
het acetyl-CoA naar buiten gebracht, waar het acetyl-CoA
wordt gebruikt. Het reducerend vermogen voor de
vetzuursynthese is NADPH afkomstig uit het PPP en uit
Citraatshuttle
de reactie malaat →pyruvaat.
Het sleutelenzym van de lipogenese is acetyl-CoA
carboxylase (ACC), deze vormt malonyl-CoA uit acetyl-
CoA. ACC wordt hormonaal en metabool gereguleerd:
Hormonaal Metabool
Insuline + (defosforyleert) Citraat +
Glucagon – (fosforyleert) AMP -
Adrenaline – (fosforyleert) Palmitaat -
Exogene lipiden in de voeding worden afgebroken tot fosfolipiden, cholesterolen, en TAG.
Enterocyten verpakken deze stoffen in lipoproteïnen: chylomicronen. Deze hebben een kern van
TAG of cholesterolester, en een mantel van een fosfolipide monolaag, cholesterol, en apoprotein
B48. De chylomicronen worden via de lymfe naar het bloed vervoerd. Hieruit komt het eerst in het
vetweefsel terecht, en enige tijd na de maaltijd (als er minder glucose is), wordt dit aangesproken.
Op de buitenkant van vetweefsel zit lipoprotein lipase, dit wordt door
insuline gestimuleerd en hecht chylomicronen, waarna het TAG afbreekt.
De vetzuren die hieruit ontstaan worden opgenomen en worden opnieuw
tot TAG gevormd. De glycerol kan hier niet worden opgenomen & gaat
terug naar de lever (gluconeogenesesubstraat). Er wordt een nieuw
glycerol gevormd uit glucose. Vetcellen hebben GLUT4, waarmee na een
maaltijd de glucose wordt opgenomen.
Het chylomicron krimpt omdat deze wordt leeggemaakt, er blijft een
‘chylomicron remnant’ over die cholesterol bevat. Dit remnant gaat terug
naar de lever, waar ze Apo E receptoren binden en terug worden
opgenomen. Dit verhaal gaat ook op voor VLDL van de lever.
Chylomicronen zijn groter dan VLDL (meer TAG). Chylomicronen zijn
aanwezig na een maaltijd; VLDL is zowel na een maaltijd als na vasten
aanwezig. VLDL heeft apoB100 en chylomicronen apoB48.
, VLDL wordt door lipogenese uit glucose of uit vetzuren uit het
vetweefsel (tijdens vasten) gemaakt. VLDL bestaat uit TAG en
cholesterol. In perifeer (vet)weefsel wordt het TAG eruit gehaald, en
houd je steeds meer cholesterol over. Het remnant dient voor het
cholesteroltransport naar weefsels. LDL is je ‘cholesterol’-pool waar
andere weefsels cholesterol uit kunnen halen.
Enkele uren na de maaltijd
zorgt glucagon voor de afgifte
van glycerol en vetzuur-
albumine complexen uit de
adipocyten.
Glucagon of adrenaline activeert een 7TM-
receptor, en activeert via cAMP en PK-A
perilipine en HS-lipase. Deze enzymen
bevorderen de TAG-afbraak.
Insuline werkt exact tegengesteld: het remt
de lipolyse door defosforylering van HS-
lipase, perilipine, en ATGL.
LDL komt uit VLDL, enkel is het verrijkt aan cholesterol &
dient het voor cholesteroltransport. Bij mensen is LDL een
risicofactor voor atherosclerose.
Perifeer bindt
LDL aan een
LDL-receptor,
waarna het
wordt ge-endocyteerd. De receptoren gaan naar het
celoppervlak terug, en de inhoud gaat naar het lysosoom.
Hier worden de cholesterolesters gehydrolyseerd en
gebruikt in het ER of opgeslagen als lipid-duppeltjes. Het
eiwitdeel wordt tot aminozuren afgebroken.
Cholesterol wordt gebruikt om het membraan soepel te
houden, in de lever om galzouten te vormen, en in veel
endocriene organen voor steroïdhormoonsynthese.
Levercellen kunnen zelf cholesterol maken uit acetyl-CoA. Als je te weinig eet, maak je het dus zelf.
Het enzym HMG-CoA reductase is de regulerende stap hierin. Normaal bij cholesterol-overschot
wordt de cholesterolsynthese via HMG-CoA reductase geremd, wordt de opslag van cholesterol als
cholesterolester (activering ACAT) geactiveerd, en wordt het aantal LDL-receptoren
gedownreguleerd op het celoppervlak.